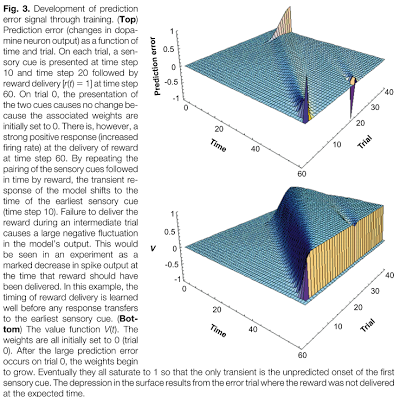
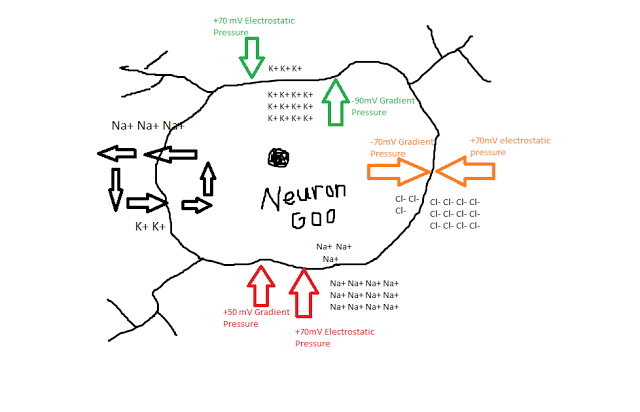
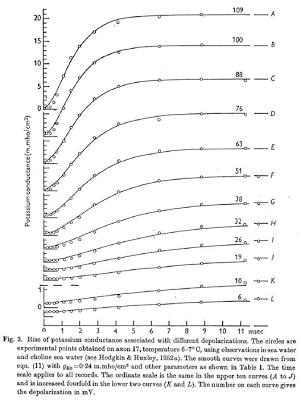
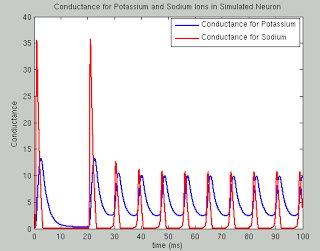
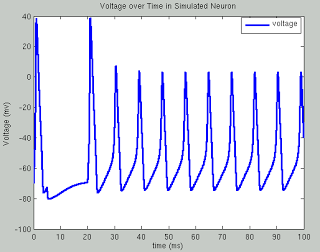
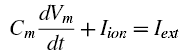In the past two weeks since I have returned from San Diego, I still find myself struck with lucid recollections during my hypopompic states, the memories of my travels twinned with hallucinations no less beautiful and no less real, were I to try distinguish the two.
When I close my eyes I can still feel the pitch-perfect weather against my skin; I can hear the purl of fountainwater underneath the lush gardens and exquisite statuary of Balboa Park; I can see those silvery, sunwarmed beaches, speckled with families and surfers and oceangoers of all stripes and ages, the terns overhead lazily riding the wind thermals and the surf below gently lapping at the shores. The man is not to be envied who does not find his spirit refreshed and invigorated by the scintillating waves of the ocean, his eros not aroused by the sight of brown-skinned beauties emerging from the sea with beads of saltwater clinging to their skin and their delicate pink toes sinking into the argentate sand; no, he is not to be envied who does not find some spark of religious awe kindled by the sight of the sun bleeding slowly into the horizon and replaced by the pale disc of the moon, pasted in that inky firmament like some ghostly wafer, overlooking the dark abyss of water out there past men's knowing, where stars are drowning and whales ferry their vast souls through the black and seamless sea.
It is well to be surrounded by such sights and sounds, as, set against the backdrop of a conference devoted to science, the conscious mind is all the more appreciative of the particulars and the practicalities of what he believes, secretly or openly, to be the healthiest, the most rigorous, the most downright of human endeavors - that of scientific inquiry. Aristotle once claimed that the twin peaks of human pleasure consist of one, sexual intercourse, and two, thinking; and once one has felt the slow-burning satisfaction of scientific experimentation, of hypotheses proposed and tested, of results surprising one in the most unexpected of ways, what right man would believe otherwise? And after a day of lively discussion and heated debate, after filling one's cup with as many poster sessions and workshops and talks as one can handle and drinking it to the lees, then one encounters the night; and, the mind still reeling from the heady fumes of science, the senses attuned to all the nuances that weren't there before, walking past the garish lighting of the restaurants and pubs of the Gaslamp district, brazen hussies with their sultry strolls and minimalist vesture calling out to each other in the darkness, steroid-inflated bouncers guarding the doors of nightclubs exuding faint rumbles of bass punctuated by shrieks and laughter - and it is here that one becomes aware of certain beauties and lusts and terrors and menace that until now were only thinly hidden. The juxtaposition of such different modes of experience makes each of them in turn that much more powerful, more savory, more piquant.
During my days at the conference inside the convention center, therefore, I expected all of the hobgoblins and ecstasies of the nights before to melt away like snow in sunshine; but even here there is an element of the surreal. Within the bowels of the convention center, several football fields long, were rows upon rows of posters, almost beyond reckoning; here is one person surrounded by intrigued colleagues, gesturing expressively with his hands, his face beaming; there is another over there who could not be more different, all alone, head down and sullenly gazing at the floor, one hand holding the opposite forearm, a perfect picture of dejection. Wind your way through the exhibition section where companies are hawking their wares, photos showing how results look before and after the application of their device, mechanical contraptions demonstrating how the latest stereotaxic equipment drills into any location without any error, no fuss, no muss. You would expect the vendors to be much more animated, to act like some sort of scientific carnival barker; but unfortunately, they sit around, this one checking her phone, that one with a saturnine expression pasted on his face and a toothpick affixed to the side of his mouth.
Upstairs through the pavilio, and enter the ballroom, where chairs are stacked in rows as neatly and ceremoniously as gravestones in a cemetery. Far away at the front of the ballroom a speaker is at the podium, a miniscule dot at this distance, but whose person is projected on several large screens hanging from the ceiling, the amount of exposure beyond the most egomaniacal totalitarian's wet dream. After a couple of hours of talks and results and diagrams of models, out the door again into the hallway, past several smaller conference rooms packed with listeners. Along my way I reach down to pick up a discarded pamphlet off the floor from the American Association for the Advancement of Science; inside, it laments that more than half of the United States population still believes in psychic phenomena such as ESP and seances. "Fifty-seven percent of American adults believe in phenomena unsupported by any evidence whatsoever," it says. "It would be better to get that number closer to zero." It then lists several resources and initiatives to educate the population to think scientifically. The younger the age at which they can stage an intervention, it seems, the better.
While I can appreciate the sentiment, part of me thinks that this feeling is misguided. I have several close colleagues who would be horrified to wake up to a world denuded of superstitions and myths; so satisfying is the sense of superiority they feel in mocking those who still hold groundless beliefs, and the repercussions so minimal, that to take that away from them would be to take away their chief joy. Conversations would dry up, bereft of the usual potent feelings of solidarity and indignation, and the only ties that used to bind them to other like-minded individuals would dissolve. No, clearly a world free of people believing crazy shit would be a catastrophe. I think that a more reasonable goal would be to get the number of persons down to about five or ten percent; that way, the Association can still claim no small measure of success in their crusade, and there will still be plenty of eccentrics left over to insult, belittle, and marginalize.
After making the rounds at all of the talks, I go back to the poster session, where new posters have been pinned up on the boards, some of them still reeking the stench of hot ink fresh off the printer. Wander around, and you begin to notice how some individuals tend to dominate the conversation surrounding a poster and poins out all the experimental flaws with a minimum of decency. To counteract this, I usually leave in one or two glaringly obvious errors in any poster I present or any paper I submit; that way, one can more easily comment on it and feel as though they have done something useful, usually leaving all of the other material alone.
That being said, however, still be aware that there is much research out there which is smoke and mirrors; having been in the game for quite some time, I can provide a short list of words and phrases that should immediately set off alarm bells in your head: neuroscience; significant; brain; rat; human; monkey; hypothesis; anterior cingulate; activation; voxels; cake; "game-changer"; default poop network. Beware the siren song of these words that charm the ear and bewitch the mind; they are beautiful but treacherous ondines who, given the chance, will wrap their briny arms around you, dragging you down to your death in the bottomless sea.
When I close my eyes I can still feel the pitch-perfect weather against my skin; I can hear the purl of fountainwater underneath the lush gardens and exquisite statuary of Balboa Park; I can see those silvery, sunwarmed beaches, speckled with families and surfers and oceangoers of all stripes and ages, the terns overhead lazily riding the wind thermals and the surf below gently lapping at the shores. The man is not to be envied who does not find his spirit refreshed and invigorated by the scintillating waves of the ocean, his eros not aroused by the sight of brown-skinned beauties emerging from the sea with beads of saltwater clinging to their skin and their delicate pink toes sinking into the argentate sand; no, he is not to be envied who does not find some spark of religious awe kindled by the sight of the sun bleeding slowly into the horizon and replaced by the pale disc of the moon, pasted in that inky firmament like some ghostly wafer, overlooking the dark abyss of water out there past men's knowing, where stars are drowning and whales ferry their vast souls through the black and seamless sea.
It is well to be surrounded by such sights and sounds, as, set against the backdrop of a conference devoted to science, the conscious mind is all the more appreciative of the particulars and the practicalities of what he believes, secretly or openly, to be the healthiest, the most rigorous, the most downright of human endeavors - that of scientific inquiry. Aristotle once claimed that the twin peaks of human pleasure consist of one, sexual intercourse, and two, thinking; and once one has felt the slow-burning satisfaction of scientific experimentation, of hypotheses proposed and tested, of results surprising one in the most unexpected of ways, what right man would believe otherwise? And after a day of lively discussion and heated debate, after filling one's cup with as many poster sessions and workshops and talks as one can handle and drinking it to the lees, then one encounters the night; and, the mind still reeling from the heady fumes of science, the senses attuned to all the nuances that weren't there before, walking past the garish lighting of the restaurants and pubs of the Gaslamp district, brazen hussies with their sultry strolls and minimalist vesture calling out to each other in the darkness, steroid-inflated bouncers guarding the doors of nightclubs exuding faint rumbles of bass punctuated by shrieks and laughter - and it is here that one becomes aware of certain beauties and lusts and terrors and menace that until now were only thinly hidden. The juxtaposition of such different modes of experience makes each of them in turn that much more powerful, more savory, more piquant.
During my days at the conference inside the convention center, therefore, I expected all of the hobgoblins and ecstasies of the nights before to melt away like snow in sunshine; but even here there is an element of the surreal. Within the bowels of the convention center, several football fields long, were rows upon rows of posters, almost beyond reckoning; here is one person surrounded by intrigued colleagues, gesturing expressively with his hands, his face beaming; there is another over there who could not be more different, all alone, head down and sullenly gazing at the floor, one hand holding the opposite forearm, a perfect picture of dejection. Wind your way through the exhibition section where companies are hawking their wares, photos showing how results look before and after the application of their device, mechanical contraptions demonstrating how the latest stereotaxic equipment drills into any location without any error, no fuss, no muss. You would expect the vendors to be much more animated, to act like some sort of scientific carnival barker; but unfortunately, they sit around, this one checking her phone, that one with a saturnine expression pasted on his face and a toothpick affixed to the side of his mouth.
Upstairs through the pavilio, and enter the ballroom, where chairs are stacked in rows as neatly and ceremoniously as gravestones in a cemetery. Far away at the front of the ballroom a speaker is at the podium, a miniscule dot at this distance, but whose person is projected on several large screens hanging from the ceiling, the amount of exposure beyond the most egomaniacal totalitarian's wet dream. After a couple of hours of talks and results and diagrams of models, out the door again into the hallway, past several smaller conference rooms packed with listeners. Along my way I reach down to pick up a discarded pamphlet off the floor from the American Association for the Advancement of Science; inside, it laments that more than half of the United States population still believes in psychic phenomena such as ESP and seances. "Fifty-seven percent of American adults believe in phenomena unsupported by any evidence whatsoever," it says. "It would be better to get that number closer to zero." It then lists several resources and initiatives to educate the population to think scientifically. The younger the age at which they can stage an intervention, it seems, the better.
While I can appreciate the sentiment, part of me thinks that this feeling is misguided. I have several close colleagues who would be horrified to wake up to a world denuded of superstitions and myths; so satisfying is the sense of superiority they feel in mocking those who still hold groundless beliefs, and the repercussions so minimal, that to take that away from them would be to take away their chief joy. Conversations would dry up, bereft of the usual potent feelings of solidarity and indignation, and the only ties that used to bind them to other like-minded individuals would dissolve. No, clearly a world free of people believing crazy shit would be a catastrophe. I think that a more reasonable goal would be to get the number of persons down to about five or ten percent; that way, the Association can still claim no small measure of success in their crusade, and there will still be plenty of eccentrics left over to insult, belittle, and marginalize.
After making the rounds at all of the talks, I go back to the poster session, where new posters have been pinned up on the boards, some of them still reeking the stench of hot ink fresh off the printer. Wander around, and you begin to notice how some individuals tend to dominate the conversation surrounding a poster and poins out all the experimental flaws with a minimum of decency. To counteract this, I usually leave in one or two glaringly obvious errors in any poster I present or any paper I submit; that way, one can more easily comment on it and feel as though they have done something useful, usually leaving all of the other material alone.
That being said, however, still be aware that there is much research out there which is smoke and mirrors; having been in the game for quite some time, I can provide a short list of words and phrases that should immediately set off alarm bells in your head: neuroscience; significant; brain; rat; human; monkey; hypothesis; anterior cingulate; activation; voxels; cake; "game-changer"; default poop network. Beware the siren song of these words that charm the ear and bewitch the mind; they are beautiful but treacherous ondines who, given the chance, will wrap their briny arms around you, dragging you down to your death in the bottomless sea.